介绍
图S1)。 慢性间歇性缺氧的模式和时间与CIH组相同。
细胞活力测定
我们使用细胞计数试剂盒8检测(CCK-8)(Beyotime,C0037,中国)来测试细胞活力。 将ROAEC添加到96孔板中,每组5个多孔板。 成型后,每孔加入10μL CCK-8溶液。
37℃孵育2h后,用酶标仪在450nm处检测吸光度。 结果按下式计算:(实验组-空白对照)/(阴性对照-空白对照)×100%。
细胞凋亡检测
CIH模型后,将细胞收集于1.5 mL ep管中,用PBS洗涤三次,然后重悬于200ul含10 µL PI和5 µL Annexin V-FITC(Beyotime,C10625,中国)的结合缓冲液中。 室温孵育10分钟后,C6流式细胞术检测细胞凋亡率。
ROS检测
通过ROS测定(Beyotime,S00335,中国)测量ROS水平。 将ROAEC种植在6孔板中。 CIH 建模后,将细胞收集到 1.5 mL ep 管中。 将 2',7'-二氯二氢荧光素二乙酸酯 (DCFH-DA) 添加到每个 ep 管中。 37℃培养箱中孵育20分钟后,用无血清DMEM洗涤细胞3次,用无血清DMEM悬浮,并进行流式细胞仪检测。
细胞内 Fe2+ 水平的检测
使用FerroOrange(F-374,同仁岛,中国)检测不同组的Fe2+水平。 将细胞收集到 1.5 mL EP 管中,用无血清 DMEM 洗涤,然后在 37°C 培养箱中用 1 µM FerroOrange 孵育 30 分钟,并通过流式细胞仪进行测试。
丙二醛 (MDA) 测定
为了评估 MDA 水平,使用了 MDA 检测试剂盒。 将细胞接种在 10 cm 培养皿中,并在建模过程后收集在 5 mL EP 管中。 根据制造商的说明,制备样品和试剂。 然后通过酶标仪测量 532 nm 和 600 nm 处的吸光度。 随后,使用标准曲线计算MDA水平。
脂质过氧化的测量
将细胞接种到 6 孔板中。 CIH造模后,将细胞收集于1.5EP管中,用无血清DMEM洗涤3次。 然后在 1 µM 4.4-二氟-5-(4-苯基-1,3-丁二烯基)-4-bora-3a,4a-二氮杂-indacene-3-十一烷酸 (C11-BODIPY 581/591) 中孵育( Thermo Fisher,美国)在 37°C 培养箱中培养 30 分钟。 无血清DMEM洗涤3次,流式细胞仪检测。
烟酰胺腺嘌呤二核苷酸(NAD+)/NADH比值检测
使用带有 WST-8 (Elabscience) 的 NADP+/NADH 测定试剂盒检测 NAD+/NADH 比率。 将细胞接种在6孔板中,用预冷的PBS洗涤,并收集在1.5 EP管中。 按照说明准备样品和试剂,然后将样品和试剂添加到标记板中。 对于 NADH 检测,样品应在 60°C 下孵育 30 分钟以去除 NAD+。 37℃孵育30分钟后,用酶标仪在450nm处检测吸光度。 使用标准曲线计算 NAD 总和 NADH。 NAD+/NADH比率=(NAD总-NADH)/NADH*100%。
代谢物分析
每组收集2*107个细胞。 离心后,将细胞保存在液氮中30s并保持在-80℃。 立即提取代谢物,并对中心碳代谢进行靶向代谢组学分析。 加入500 μL预冷的MeOH/H2O(3/1,v/v)后,将样品在干冰中预冷,并在液氮中重复冻融3次; 收集 400 μL 等份的澄清上清液并通过旋转干燥。 然后,用200μL纯化水复溶样品。 重构的样品在通过离心管过滤器过滤之前进行涡旋,随后转移到注射瓶中的插入物中以进行 HPIC-MS/MS 分析。 使用配备 Dionex IonPac AS11-HC (2×250 mm) 和 AG11-HC (2 mm×50 mm) 色谱柱的 Thermo Scientific Dionex ICS-6000 HPIC 系统(Thermo Scientific)进行 HPIC 分离。 采用配备电喷雾电离 (ESI) 接口的 AB SCIEX 6500 QTRAP+ 三重四极杆质谱仪 (AB Sciex) 进行检测开发。 使用热图R包(corrplot)(版本0.89)来分析中心碳代谢的热图。
实时荧光定量PCR
使用总RNA分离试剂盒V2(Vazyme,中国)提取RNA。 使用 HiScript III All-in-one RT SuperMix Perfect for qPCR(Vazyme,R333-01,中国)对 cDNA 进行反转录。 RT-qPCR 在 (ABI QuantStudio 5) ABI 7500 热循环仪(Applied Biosystems, Foster City, CA, USA)上使用 SYBR Green PCR Master Mix(Vazyme,Q712,中国)进行。 检测到 GPX4 和 SLC7A11。 表格1 显示适当的引物。 通过2-Δ(CT)法计算mRNA的相对表达水平。
表格1 PCR引物序列
蛋白质印迹法
使用放射免疫沉淀分析(RIPA)裂解缓冲液(Beyotime,中国)提取蛋白质。 采用BCA蛋白浓度试剂盒(Beyotime,中国)检测蛋白浓度。 然后进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)(美伦生物,中国)分离蛋白质。 电泳将蛋白质分子转移到膜上(Millipore, 150 Billerica, MA, USA)。 然后,将膜浸泡在5%脱脂奶粉中,并溶解在Tris Buffer Solution Tween(TBST)(servicebio,G0001-2L,中国)中2小时。 一抗 SLC7A11(溶质载体家族 7 成员 11,Abcam,ab175186,1:1000)、GPX4(谷胱甘肽过氧化物酶 4,Abcam,ab252833,1:1000)和 β-肌动蛋白(Abcam,ab5694,1:1000)然后在 4°C 下孵育过夜。 TBST洗膜后,与羊抗兔IgG二抗(Abcam,ab205718,1:10,000)室温孵育1 h,然后用TBST洗3次。 一些印迹在浸泡在抗体中之前被分开。 借助改进的ECL试剂盒(Meilunbio,MA0186-1,中国),图像由Image J拍摄和分析。
透射电子显微镜
造模后弃去培养基,加入电镜固定液,4℃培养2~4h,低速离心后收集细胞。 将样品包埋在 1% 琼脂糖中,并用 0.1m 磷酸盐缓冲液冲洗三次,每次 15 分钟。 然后在室温下将它们固定在 1% 锇酸和 0.1m 磷酸盐缓冲液混合物中。 然后用0.1m磷酸盐缓冲液冲洗三次,每次15分钟。 样品依次用30-50-70-80-95-100-100%乙醇室温脱水15 min,100%丙酮室温脱水两次,每次15 min。 包埋后,将超薄切片切成70nm,用柠檬酸铅染色,并用透射电子显微镜(Hitachi,HT7700,日本)拍照。
线粒体膜电位的检测
JC-1法(Beyotime,C2006,中国)用于测试线粒体膜电位(MMP)。 按照说明书配制JC-1工作液,每组加入500 μL,混匀,37℃孵育20 min。 300g/min离心5分钟后,滴去上清,用预冷的1*JC-1 Assay Buffer洗涤两次。 用1*JC-1 Assay Buffer重悬细胞,然后用流式细胞仪检测。
统计分析
结果全部显示为平均值±标准差。 使用GraphPad Prism 7.0(GraphPad Software Inc.,美国)进行统计分析。 学生-t两组间采用-检验进行统计学分析,P<0.05为有统计学意义。
结果
CIH 诱导的 ROAEC 损伤和铁死亡
CIH条件下暴露36 h后,检测细胞凋亡率、ROS水平、Fe2+水平、脂质ROS水平、MDA水平(图1A-D,图1H)与对照组相比显着增加(p < 0.05)。 CIH 暴露 36 小时后,ROAEC 的细胞活力和 NAD+/NADH 比率显着降低(图1E 和 我)(p < 0.05)。 与对照组相比,CIH组GPX4和SLC7A11的mRNA和蛋白水平显着下降,均p < 0.05,如图所示 图1F-G。 原始蛋白质印迹图像如图 S2 所示。

图1 CIH 诱导的 ROAEC 损伤和铁死亡。 (A)、CIH组和对照组的细胞凋亡率; (乙)、CIH组和对照组ROS水平; (C)、CIH组和对照组的Fe2+水平; (D)、CIH组和对照组脂质ROS水平; (乙),CIH组和对照组ROAEC细胞活力下降; (F)、CIH组和对照组GPX4、SLC7A11 mRNA水平; (G)、CIH组和对照组GPX4和SLC7A11蛋白水平; (H)、CIH组和对照组的MDA水平; (我)、CIH组和对照组的NAD+/NADH比值。 数据重复至少三次并显示为平均值±SD。 *p<0.05; **p < 0. 01; ***p < 0. 001; ****p < 0.0001.
Fer-1 减轻 CIH 相关的 ROAEC 损伤和铁死亡
CIH和Fer-1联合处理后,细胞凋亡率、ROS水平、Fe2+水平、脂质ROS水平、MDA水平(图2A-D,图2H)减少; 而细胞活力和NAD+/NADH比值较CIH组有所增加(图2E、图2I)(所有 p < 0.05)。 在CIH+Fer-1组中,与CIH组相比,GPX4和SLC7A11的mRNA和蛋白水平增加(图2F-G)(均 p < 0.05)。 原始蛋白质印迹图像如图 S3 所示。

Fer-1 减轻 CIH 诱导的线粒体结构和功能损伤
为了观察线粒体的结构和功能,我们进行了TEM和MMP检测。 TEM结果显示,与对照组相比,CIH组线粒体膜受损,并出现基质溢出现象(图3A)。 JC-1检测结果显示,CIH组MMP低于对照组(图3B)。 与Fer-1共同治疗后,CIH引起的线粒体结构和MMP损伤被逆转(图3C-D)。
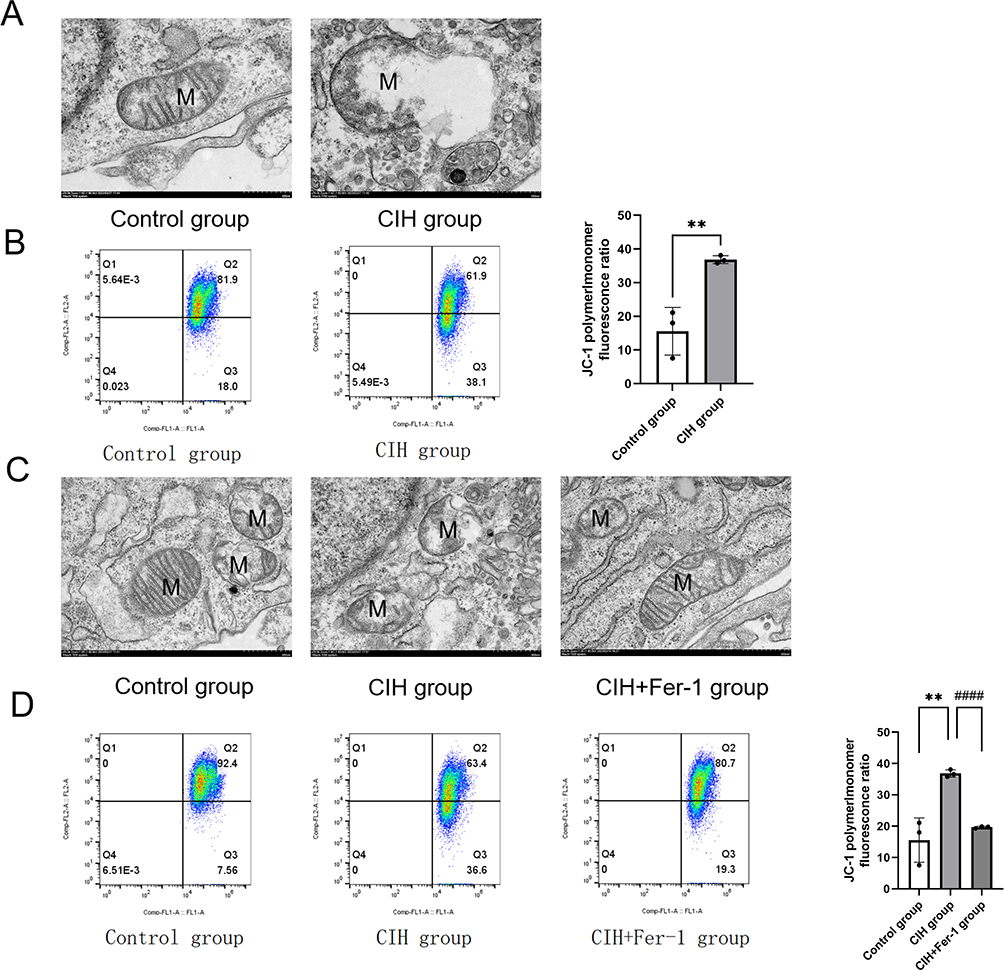
图3 Fer-1 减轻了 CIH 治疗的 ROAEC 中的线粒体损伤。 (A),对照组和CIH组的透射电镜(TEM)照片; (乙),对照组和CIH组线粒体膜电位(MMP)水平。 (C)、对照组、CIH组、CIH+Fer-1组的TEM照片。 (D)、对照组、CIH组、CIH+Fer-1组的MMP水平。 数据显示为平均值±SD。 与对照组相比,**p < 0. 01; 与 CIH 组相比,####p < 0. 0001。
Fer-1 缓解 CIH 诱导的线粒体代谢紊乱

图4 Fer-1 改善了 CIH 诱导的 TCA 循环代谢异常。 对照组和 CIH 组中中心碳代谢显着调节代谢物的层次聚类(n = 6,生物重复)。(A) 色标表示相对代谢物水平:红色表示上调,蓝色表示下调。 (乙), TCA 循环中代谢物改变的示意图。(C),对照组、CIH组和CIH+Fer-1组中中心碳代谢显着调节的代谢物的层次聚类(n = 6,生物重复)。 色标表示相对代谢水平:红色表示上调,蓝色表示下调。 (D),CIH 增加了 TCA 循环中代谢物的水平。 Fer-1治疗减轻了CIH引起的代谢物的增加。 数据显示为平均值±SD。 与对照组相比,**p < 0. 01,***p < 0. 001,****p < 0. 0001; 与 CIH 组相比,### p < 0. 001,####p < 0. 0001。
三羧酸(TCA)循环是位于线粒体基质中的酶途径。 筛选出TCA循环中差异表达的关键代谢物,如图所示 图4B。 比较各组中α-酮戊二酸(αKG)、琥珀酸、富马酸、苹果酸、2-氧代丁酸和顺乌头酸的表达水平。 CIH 暴露增加了 TCA 循环中的代谢物水平。 与 CIH 组相比,Fer-1+CIH 组的 TCA 循环中的代谢物水平降低,如图所示 图4D (所有 p < 0.05)。
讨论
目前的研究表明,CIH 暴露会导致 ROAEC 铁死亡。 铁死亡抑制剂Fer-1可有效抑制这种损伤。 此外,在 CIH 处理的 ROAEC 中,线粒体形态、功能和 TCA 循环中的代谢物发生了变化,而 Fer-1 可以逆转这种变化。
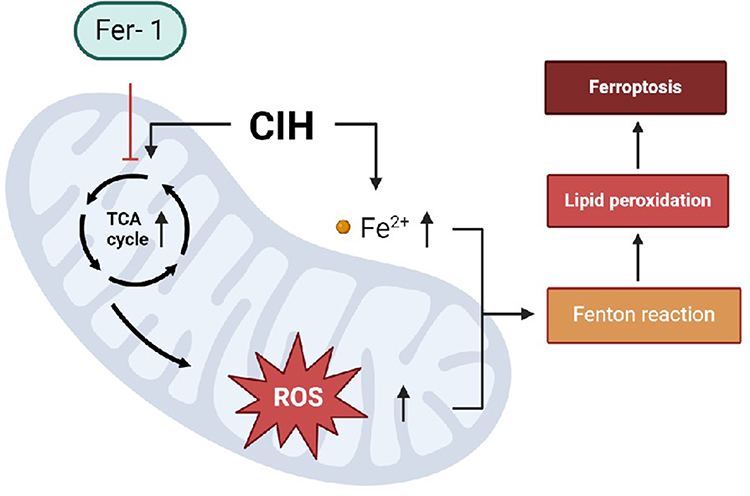
图5 Fer-1 逆转 CIH 诱导的主动脉内皮细胞铁死亡的可能分子机制。 在ROAEC中,CIH处理后,TCA循环中重要代谢产物的活性增加,导致脂质ROS的积累、线粒体膜电位的变化以及线粒体结构的破坏,最终导致铁死亡。 与 Fer-1 共同治疗可通过重新编程 TCA 循环的代谢物和线粒体功能来逆转铁死亡。
这项研究存在一些局限性。 该研究仅限于细胞实验。 体外和体内研究的结果可能存在一些差异。 因此,我们今后将通过动物实验来证实Fer-1在CIH相关心血管损伤中的作用。
结论
CIH 诱导的 ROAEC 铁死亡。 Fer-1逆转了TCA代谢物的水平,然后减少了ROS的积累和线粒体结构的破坏,最终逆转了ROAEC中CIH诱导的铁死亡的发生。
缩写
OSA,阻塞性睡眠呼吸暂停; CIH,慢性间歇性缺氧; Fer-1,铁他汀-1; ROAEC,大鼠动脉内皮细胞; SLC7A11,胱氨酸/谷氨酸逆向转运蛋白溶质载体家族7成员11; GPX4、谷胱甘肽过氧化物酶4; CCK-8,细胞计数试剂盒-8; ROS,活性氧; COPD,慢性阻塞性肺疾病; DMEM,杜尔贝科改良伊格尔培养基; DCFH-DA,2',7'-二氯二氢荧光素二乙酸酯; TEM、透射电子显微镜; RIPA,放射免疫沉淀分析; SDS-PAGE,十二烷基硫酸钠-聚丙烯酰胺凝胶电泳; TBST,Tris 缓冲液吐温; PBS,磷酸盐缓冲盐水; TCA,三羧酸循环; MMP,线粒体膜电位; αKG,α-酮戊二酸; NAD+,烟酰胺腺嘌呤二核苷酸; MDA、丙二醛; C11-BODIPY 581/591,4,4-二氟-5-(4-苯基-1,3-丁二烯基)-4-bora-3a,4a-二氮杂-indacene-3-十一烷酸; PPP,戊糖磷酸途径。
数据共享声明
本研究使用的数据可通过通讯作者获得。
资金
该研究得到了福建省自然科学基金项目(编号:2023J01554)、福建省财政联合基金项目(编号:BPB-LNF2021)、国家自然科学基金项目(编号:82170101)的资助。
披露
作者报告在这项工作中没有利益冲突。
参考
1. Gottlieb DJ,新墨西哥州旁遮普语。 阻塞性睡眠呼吸暂停的诊断和治疗:综述。 美国医学会杂志。 2020;323(14):1389–1400。 doi:10.1001/jama.2020.3514
2. Drager LF、Togeiro SM、Polotsky VY 等。 阻塞性睡眠呼吸暂停:肥胖和代谢综合征的心脏代谢风险。 J Am Coll Cardiol 杂志, 2013;62(7):569–576。 doi:10.1016/j.jacc.2013.05.045
3. Chen L,Zou S,Wang J。阻塞性睡眠呼吸暂停综合征(OSA/OSAHS)与冠状动脉粥样硬化风险的关联:系统评价和荟萃分析。 医学计算数学方法。 2022;2022:8905736。 号码:10.1155/2022/8905736
4. Badran M、Ayas N、Laher I。睡眠呼吸暂停的心血管并发症:氧化应激的作用。 氧化物与电池寿命。 2014;2014:985258。 号码:10.1155/2014/985258
5. Badran M,Gozal D. PAI-1:阻塞性睡眠呼吸暂停血管功能障碍的主要参与者?。 国际分子科学杂志。 2022;23(10):5516。 doi:10.3390/ijms23105516
6. 迪卡洛夫 SI,纳扎雷维奇 RR。 血管紧张素 II 诱导线粒体活性氧的产生:潜在机制及其与心血管疾病的相关性。 抗氧化氧化还原信号。 2013;19(10):1085–1094。 doi:10.1089/ars.2012.4604
7. Puddu P、Puddu GM、Galletti L 等。 线粒体功能障碍作为动脉粥样硬化形成的起始事件:一个合理的假设。 心脏病学。 2005;103(3):137–141。 号码:10.1159/000083440
8. 赵丽,刘涛,窦子杰,等。 CB1 受体拮抗剂利莫那班可预防大鼠慢性间歇性缺氧引起的肾损伤。 BMC肾病。 2021;22(1):153。 号码:10.1186/s12882-021-02362-6
9. 宋JQ,蒋丽莲,付CP,等。 杂合子 SOD2 缺失通过 mtROS-NLRP3 信号通路恶化慢性间歇性缺氧诱导的肺部炎症和血管重塑。 药理学学报。 2020;41(9):1197–1207。 doi:10.1038/s41401-019-0349-y
10. 派 PY,林 YY,于 SH,等。 血管紧张素 II 受体阻滞剂厄贝沙坦可减弱睡眠呼吸暂停诱导的心脏细胞凋亡,并增强心脏存活率和 Sirtuin 1 上调。 睡眠呼吸。 2022;26(3):1161–1172。 doi:10.1007/s11325-021-02499-6
11. Shi C,Guo H,Liu X。血小板线粒体移植可挽救缺氧/复氧诱导的线粒体功能障碍和涉及 FUNDC2/PIP3/Akt/FOXO3a 轴的神经元细胞死亡。 细胞移植。 2021;30:9636897211024210。 号码:10.1177/09636897211024210
12. 陈杰,朱浩,陈Q,等。 铁死亡在慢性间歇性缺氧引起的肺损伤中的作用。 BMC 普尔医学。 2022;221:488。 BMC 肺科。 10.1186/s12890-022-02262-x
13. Arnaud C、Billoir E、De Melo Junior AF 等。 慢性间歇性缺氧引起的心血管和肾功能障碍:从适应到适应不良。 生理学杂志。 2023;601(24):5553–5577。 doi:10.1113/JP284166
14. Dixon SJ、Lemberg KM、Lamprecht MR 等人。 铁死亡:一种铁依赖性非凋亡细胞死亡形式。 细胞。 2012;149(5):1060–1072。 doi:10.1016/j.cell.2012.03.042
15. 李杰,曹芳,尹红莲,等。 铁死亡:过去、现在和未来。 细胞死亡疾病。 2020;11(2):88。 doi:10.1038/s41419-020-2298-2
16. 赵文康,周永,徐天涛,等。 铁死亡:心肌缺血再灌注损伤的机遇与挑战。 氧化物与电池寿命。 2021;2021:9929687。 号码:10.1155/2021/9929687
17. 李X,马宁,徐J,等。 靶向铁死亡:缺血再灌注损伤的病理机制和治疗。 氧化物与电池寿命。 2021;2021:1587922。 号码:10.1155/2021/1587922
18. 陈立德,吴瑞华,黄YZ,等。 铁死亡在大鼠慢性间歇性缺氧性肝损伤中的作用。 睡眠呼吸。 2020;24(4):1767–1773。 doi:10.1007/s11325-020-02091-4
19. Dan Dunn J、Alvarez LA、Zhang X 等人。 活性氧和线粒体:细胞稳态的联系。 氧化还原生物。 2015;6:472–485。 doi:10.1016/j.redox.2015.09.005
20. Paul BT、Manz DH、Torti FM 等。 线粒体和铁:当前的问题。 血液专家修订版。 2017;10(1):65–79。 号码:10.1080/17474086.2016.1268047
21. Stockwell BR、Friedmann Angeli JP、Bayir H 等人。 铁死亡:一种连接新陈代谢、氧化还原生物学和疾病的受调节细胞死亡关系。 细胞。 2017;171(2):273–285。 doi:10.1016/j.cell.2017.09.021
22. 高明,姜旭。吃还是不吃——铁死亡的代谢味道。 当前细胞生物学观点。 2018;51:58–64。 doi:10.1016/j.ceb.2017.11.001
23. Jiang X, Stockwell BR, Conrad M. 铁死亡:机制、生物学和在疾病中的作用。 Nat Rev Mol 细胞生物学。 2021;22(4):266–282。 号码:10.1038/s41580-020-00324-8
24. 唐东,陈X,康R,等。 铁死亡:分子机制和健康影响。 细胞研究。 2021;31(2):107–125。 doi:10.1038/s41422-020-00441-1
25. 孙Y,Berleth N,吴W,等。 Fin56 诱导的铁死亡得到自噬介导的 GPX4 降解的支持,并与 mTOR 抑制协同作用以杀死膀胱癌细胞。 细胞死亡疾病。 2021;12(11):1028。 doi:10.1038/s41419-021-04306-2
26. Chen D、Tavana O、Chu B 等人。 NRF2 是 ARF 在不依赖于 p53 的肿瘤抑制中的主要靶标。 分子细胞。 2017;68(1):224–32.e4。 doi:10.1016/j.molcel.2017.09.009
27. 罗明,王永志,痛风普文. x(c)-胱氨酸/谷氨酸逆向转运蛋白:癌症和其他疾病治疗的潜在靶点。 细胞生理学杂志。 2008;215(3):593–602。 doi:10.1002/jcp.21366
28. Javaheri S、Barbe F、Campos-Rodriguez F 等人。 睡眠呼吸暂停:类型、机制和临床心血管后果。 J Am Coll Cardiol 杂志, 2017;69(7):841–858。 doi:10.1016/j.jacc.2016.11.069
29. 高明,易军,朱军,等。 线粒体在铁死亡中的作用。 分子细胞。 2019;73(2):354–63.e3。 doi:10.1016/j.molcel.2018.10.042
30. 李春,张勇,刘杰,等。 线粒体 DNA 应激触发自噬依赖性铁死亡。 自噬。 2021;17(4):948–960。 号码:10.1080/15548627.2020.1739447
31. Lee H、Zandkarimi F、张 Y 等。 能量应激介导的 AMPK 激活可抑制铁死亡。 自然细胞生物学。 2020;22(2):225–234。 doi:10.1038/s41556-020-0461-8
32. Na YR,Je S,Seok SH。 炎症性疾病和癌症中巨噬细胞的代谢特征。 癌症快报。 2018;413:46–58。 doi:10.1016/j.canlet.2017.10.044
33. Bailey JD、Diotallevi M、Nicol T 等人。 一氧化氮通过 TCA 循环调节和衣康酸积累来调节炎症巨噬细胞的代谢重塑。 细胞代表。 2019;28(1):218–30.e7。 doi:10.1016/j.celrep.2019.06.018
34. 刘平,吴东,段健,等。 NRF2 通过 FOCAD-FAK 信号通路调节人 NSCLC 细胞对胱氨酸剥夺诱导的铁死亡的敏感性。 氧化还原生物。 2020;37:101702。 doi:10.1016/j.redox.2020.101702
35. 罗Y,陈鹏,杨L,等。 3,4-二羟基苯甲醛对脑缺血再灌注损伤脑保护作用的代谢组学分析及药理学验证。 带绳子的鼹鼠。 2023;27(1):9。
1713841889
2024-04-23 02:21:44
#CIH诱导大鼠动脉内皮细胞铁死亡