
介绍
乳腺癌对女性的生命和健康构成严重威胁。1 随着生活方式和饮食习惯的改变以及生育率的提高,乳腺癌的发病率逐年上升,已成为当今的热点问题。2,3 乳腺癌是一种复杂且异质的疾病。 随着靶向治疗技术的应用,乳腺癌患者的临床疗效得到了极大的提高。4 然而,仍然缺乏高度特异性的靶向标记。 因此,积极探索乳腺癌防治的潜在分子靶点、改善患者预后是当今乳腺癌研究的重点课题。 乳腺癌的恶性进展是一个涉及多因素变化、多基因调控和多信号通路的复杂过程。5 深入探索乳腺癌恶性进展的调控机制,有望使乳腺癌患者的临床治疗更加精准化、个性化。
我们选择LINC02418作为研究靶点,探索乳腺癌患者治疗和预后的潜在生物标志物。
材料和方法
临床标本
这项研究是回顾性的。 2016年至2018年,内江市第一人民医院共有129名符合以下标准的乳腺癌患者。
纳入标准:
排除标准:
收集129例受试者切除的乳腺癌组织和相应的正常癌旁组织样本,用液氮快速冷冻,-80℃保存。 所有样本均由两名专业病理学家诊断为乳腺癌。 所有受试者均知情同意,本研究获得内江市第一人民医院伦理委员会授权(编号201589)。 该研究符合赫尔辛基宣言。
细胞培养
正常乳腺上皮细胞MCF-10A和乳腺癌细胞系SK-BR-3、MCF-7、MDA-MB-231、MDA-MB-468购自中国科学院上海细胞库。 实验中,首先将细胞解冻,然后重悬于含有+1%青霉素-链霉素溶液+10%胎牛血清的DMEM培养基中。 将细胞转移至培养皿中并在37°C和5%CO2下观察。
细胞转染
选择对数生长且状态良好的细胞作为转染对象。 将Lipofectamine 2000(Invitrogen,美国)与敲低LINC02418片段(si-LINC02418)、阴性对照片段(si-NC)、miR-766-5p抑制剂(miR-inhibitor)、miR-766-5p模拟物混合获得转染复合物(miR-mimic)、miRNA 阴性对照片段 (miR-NC)、带有 miR 抑制剂的 si-LINC02418 (si+miR-inhibitor) 及其阴性对照 (si+miR-NC)(序列显示于表S1)。 转染期间,将细胞悬液和转染复合物添加到孔板中并充分混合。 在相同环境下继续培养48h,用于后续实验。
双荧光素酶报告基因检测
将含有miR-766-5p结合位点的LINC02418序列和含有突变位点的LINC02418序列分别称为WT-LINC02418和MT-LINC02418,将其克隆至pGL3荧光素酶载体中,构建重组载体。 然后将上述载体分别与miR-NC、miR-mimic和miR-inhibitor转染复合物共转染至乳腺癌细胞中。 培养48h后,检测转染细胞中LINC02418荧光素酶的活性。
定量PCR
为了检测LINC02418和miR-766-5p的表达水平,需要先通过Trizol法(Invitrogen,美国)提取总RNA,然后通过PrimeScript RT Enzyme Mix I(TaKaRa,日本)试剂盒,由 TB Green Premix Ex Taq II 试剂盒(TaKaRa,日本)扩增。 使用 2−ΔΔct 方法,以 GAPDH 和 U6 分别作为 LINC02418 和 miR-766-5p 的内标,计算扩增序列的相对表达量。 引物序列如表S2所示。
CCK8检测
取生长细胞的对数,重悬并计数,接种至96孔板中并置于培养箱中进一步培养。 培养后24、48、72和96小时加入CCK8试剂,然后继续孵育2小时。 采用酶标仪(Thermofisher,美国)测定各组细胞在450nm波长处的吸光度(OD450)。
Transwell实验
对于迁移实验,将转染后的细胞消化,然后重悬于空白DEME培养基中。 取一定量的细胞悬液接种于Transwell上室,下室加入含血清的DMEM培养基,整个过程避免气泡。 侵袭实验需要用基质胶覆盖Transwell上室,其他步骤与迁移实验相同。 在培养箱中孵育24小时,然后取出小室进行固定和染色。 最后清洗干燥后,在显微镜下观察计数。
统计分析
采用SPSS 26.0和GraphPad Prism 7.0软件对相关数据进行分析。 采用 T 检验和卡方检验分别检查组间差异以及 LINC02418 与临床病理变量之间的关系。 Kaplan-Meier 和多变量 Cox 回归分析评估了 LINC02418 对乳腺癌患者的预后能力。 通过Spearman相关分析分析LINC02418与miR-766-5p之间的关系。 本研究中的所有实验均独立进行并重复3次,以平均值±标准差表示。 磷 < 0.05 被认为具有统计显着性。
结果
LINC02418在乳腺癌中的表达及临床意义
LINC02418 表达在癌症组织中上调(图1a)和癌细胞(图1b),与正常乳腺组织和细胞相比。 根据乳腺癌组织中 LINC02418 的平均表达情况,129 名患者被分为两组:低 LINC02418 组 61 名,高 LINC02418 组 68 名。 如图所示 表格1LINC02418高表达和低表达患者的TNM分期(p=0.003)和淋巴结转移(p=0.007)存在显着差异。 LINC02418(r=2.951, p=0.012),淋巴结转移(r=0.417, p=0.015) 和 TNM 阶段 (r=0.408, p= 0.031)可作为乳腺癌患者预后的独立因素(图1c)。 高 LINC02418 组患者的 5 年生存率低于低 LINC02418 组(对数秩 p=0.045, 图1d)。
表格1 LINC02418与患者临床病理特征的关联
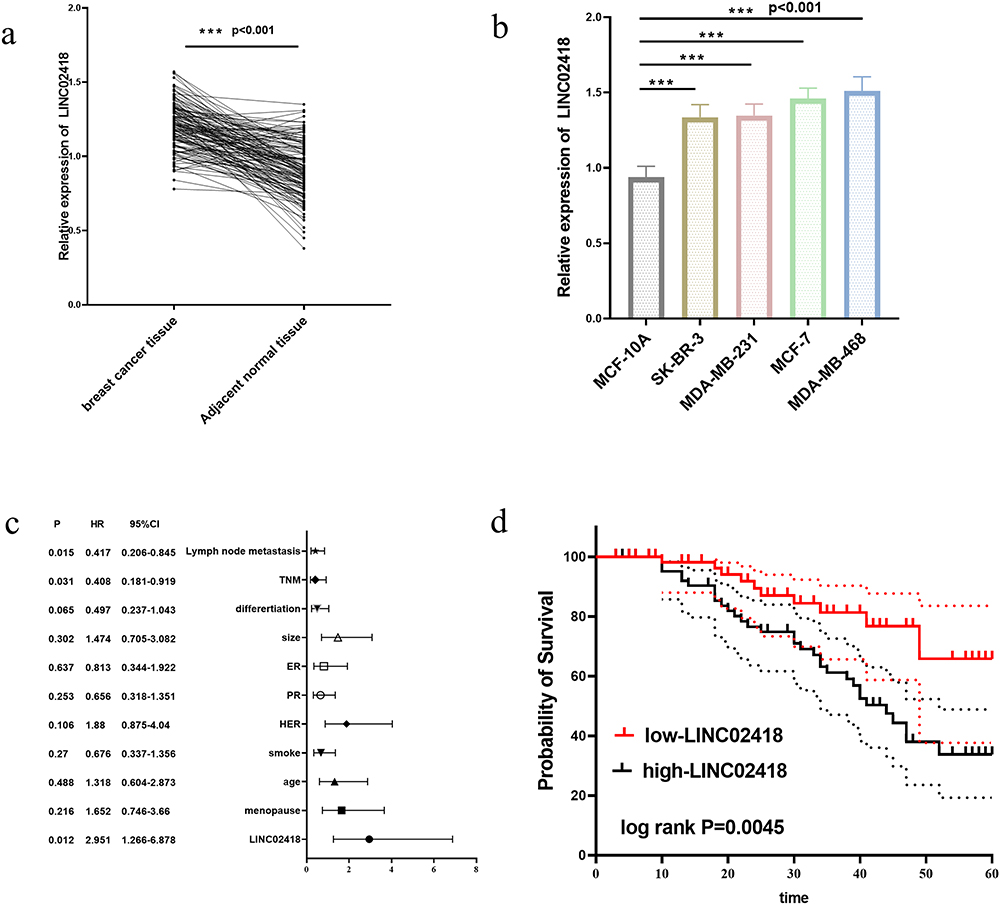
图1 乳腺癌组织中 LINC02418 的表达量高于邻近正常组织(A)。 与正常乳腺细胞 (MCF-10A) 相比,乳腺癌细胞中的 LINC02418 表达上调(乙)。 影响患者预后的独立因素的Cox多因素分析(C)。 LINC02418高表达组5年生存率低(d)。 ***p<0.001,与正常组织和正常细胞相比。
LINC02418 与 miR-766-5p 在乳腺癌中的相互作用
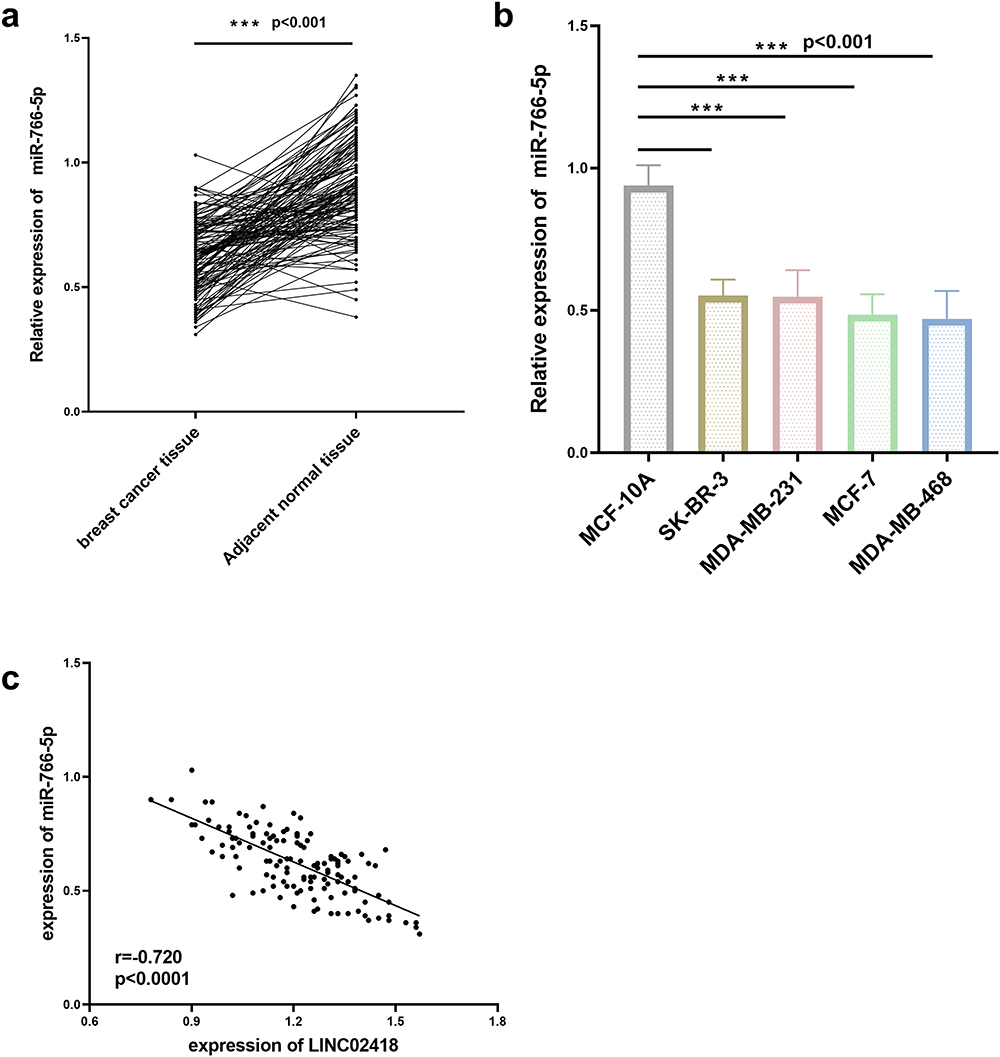

图3 miR-766-5p 影响 LINC02418 荧光素酶活性(A)。 si-LINC02418 有效敲低 LINC02418 表达(乙)。 LINC02418 的敲低会降低 miR-766-5p 的表达(C)。 **p与对照组相比,< 0.001。 ##p与 si-LINC02418 组相比 < 0.01。
缩写:NC:阴性对照。 si:沉默,LINC02418 表达敲低。
LINC02418/miR-766-5p对乳腺癌细胞活性的影响
为了研究LINC02418/miR-766-5p对乳腺癌细胞活性(包括增殖、迁移和侵袭)的影响,分别进行了CCK8和Transwell实验。 结果显示,转染si-LINC02418的乳腺癌细胞的MCF-7和MDA-MB-468的增殖率均较对照组显着降低,而转染miR-766-5p抑制剂可逆转这一现象。图4a)。 同样,迁移(图4b)和入侵(图4c转染si-LINC02418和miR-766-5p抑制剂后两种乳腺癌细胞的增殖趋势与癌细胞的增殖趋势一致。

图4 LINC02418 的敲低抑制了细胞不良的生物学行为,而 miR-766-5p 的低表达可以逆转这种行为。交流电)。 **p与对照组相比<0.001; ##p与 si-LINC02418 组相比 < 0.01。
讨论
乳腺癌的高患病率引起了女性的高度关注。17 号,18 乳腺癌发病机制复杂,发病率高,预后差,严重影响女性健康生活。19,20 随着lncRNA研究的深入,越来越多的lncRNA被发现在乳腺癌中发挥重要作用。21,22 据报道,IncRNA UCA1 通过吸附 miR-206 上调乳腺癌细胞中的蛋白酪氨酸磷酸酶 PTP1B,从而增强细胞增殖。23 LncRNA NEAT1通过竞争性吸附miR-448促进EMT关键转录因子ZEB1的表达,进而加速乳腺癌细胞的侵袭和转移。24 lncRNA TINCR可以靶向miR-125b,促进HER2+乳腺癌细胞的EMT转移,并增加癌细胞对曲妥珠单抗的耐药性,是乳腺癌潜在的分子标志物和治疗靶点。25 可见乳腺癌中有很多表达失调的lncRNA。 探索新型lncRNA在乳腺癌中的特殊作用和生物学功能,探讨其调控关系和分子机制,可为乳腺癌的临床靶向治疗提供理论基础。
LncRNA有多种分类系统,其中长基因间ncRNA(LINCRNA)位于两个蛋白质编码基因之间,可以独立转录。 已发现多种 LINCRNA 参与乳腺癌的发生。 LINC00665被发现通过负向调节miR-3619-5p发挥促癌作用。26 研究发现LINC01094在乳腺癌中表达上调,可作为预后不良的标志物。27 同样,本研究发现LINC02418高表达组和低表达组的淋巴结转移和TNM分期存在显着差异,并且LINC02418高表达组乳腺癌患者的死亡率更高。 因此,推测LINC02418有潜力成为疾病恶化和预后不良的生物标志物。 LINC02418在肺腺癌细胞中高表达,敲低其表达可以抑制癌细胞的活性。 LINC02418通过靶向miR-4677-3p促进着丝粒支架1(KNL1)的表达,从而促进肺癌细胞的发育。15 已知LINC02418表达上调,LINC02418通过海绵miR-34b-5p影响BCL2信号通路,从而增强结肠癌组织的生命活性。16 同样,LINC02418在非小细胞肺癌的组织和细胞中均上调,而LINC02418的过表达可显着增强非小细胞肺癌的增殖和侵袭活性并抑制细胞凋亡。28 据此,我们得出结论:LINC02418可能参与乳腺癌的发生和进展。 对其进行了进一步的研究。 研究发现LINC02418可以促进乳腺癌细胞的活性,发挥癌基因的作用。
通过靶向 miR-4677-3p,LINC02418 导致 SEC61G 表达上调并促进非小癌细胞的活性。28 LINC02418通过靶向miR-372-3p间接调控EPHA2,为降低结直肠癌细胞耐药性、促进结直肠癌的治疗奠定重要基础。12 然而,lncRNA在乳腺癌中的分子调控机制仍不清楚,因此本研究深入探讨关键lncRNA在乳腺癌进展中的分子调控。 荧光素酶活性报告表明 LINC02418 和 miR-766-5p 之间有一个靶向结合位点。 此外,LINC02418被发现负向调节miR-766-5p表达。 miR-766-5p抑制剂可以逆转LINC02418低表达对癌细胞生物学行为的抑制作用。 在三阴性乳腺癌 (TNBC) 中,发现 miR-766-5p 通过与 yes 相关蛋白 1 (YAP1) 结合来调节癌细胞的发育。29 此外,miR-766-5p参与多种癌症的发生、发展并发挥关键作用。 例如,miR-766-5p通过靶向癌基因激肽释放肽相关肽酶12(KLK12)参与肺癌发生的分子机制,CASC15通过负向调节miR-766-5p间接促进癌症的发生。 5点。30 miR-766-5p 在甲状腺乳头状细胞的上皮间质转化中发挥重要作用,LCC-MPEG1-1 通过海绵 miR-766-5p 促进癌症发展。31 因此推测LINC02418通过负向调节miR-766-5p来促进乳腺癌细胞的增殖和转移活性。 这一结论将在后续的体内动物实验中得到进一步验证。
乳腺癌由三种亚型组成:ER+ Luminal、ERBB2+、ER+ Luminal 和 ER-/PR-/ERBB2- 三阴性乳腺癌 (TNBC)。 不同亚型乳腺癌患者的发展及预后存在一定差异,因此推测LINC02418在不同亚型乳腺癌中的表达及临床意义不同。 本研究并未探讨LINC02418在不同亚型中的作用,将在未来的研究中进一步深化,旨在推进LINC02418在临床实践中的使用。
结论
总之,LINC02418在乳腺癌中表达上调,有可能成为乳腺癌不良预后的生物标志物。 LINC02418 可能通过 miR-766-5p 的负调控参与癌细胞增殖和转移。
披露
作者报告在这项工作中没有利益冲突。
参考
1. Yuanliang Yan QL,Liu Y,Zhou S,Zhijie X。COL12A1作为预后生物标志物与乳腺癌的免疫治疗反应相关。 内分泌相关癌症。 2023;30。
2. Soofiani SR、Hosseini K、Ebrahimi T 等人。 mir-126 在乳腺癌中的预后价值和生物学作用。 微小RNA。 2022;11(2):95–103。 号码:10.2174/1876402914666220428123203
3. 杨X,唐涛,周涛。化生性乳腺癌的预后和临床病理特征:荟萃分析。 药品。 2022;101(49):e32226。 号码:10.1097/MD.0000000000032226
4. 苗 H、哈特曼 M、Bhoo-Pathy N 等。 预测亚洲女性新发转移性乳腺癌的生存:系统评价和验证研究。 公共图书馆一号。 2014;9(4):e93755。 doi:10.1371/journal.pone.0093755
5. Chen H, Xie G, Luo Q, Yang Y, Hu S. 乳腺癌细胞周期进展中的调控 miRNA、circRNA 和 lncRNA。 功能整合基因组学。 2023;23(3):233。 doi:10.1007/s10142-023-01130-z
6. 姚ZT,杨YM,孙MM,等。 关于癌症中长链非编码 RNA 和 RNA 结合蛋白之间相互作用的新见解。 癌症通讯。 2022;42(2):117–140。 doi:10.1002/cac2.12254
7. 谭YT,林JF,李T,李JJ,徐RH,鞠HQ。 LncRNA介导的癌症能量代谢的翻译后修饰和重编程。 癌症通讯。 2021;41(2):109–120。 doi:10.1002/cac2.12108
8. Mas AM、Huarte M。作为癌症生物标志物的长非编码 RNA 特征。 J 临床肿瘤学。 2023;41(16):3059–3062。 号码:10.1200/JCO.23.00381
9. 孔X,段Y,桑Y,等。 LncRNA-CDC6 促进乳腺癌进展,并作为 ceRNA 通过海绵 microRNA-215 来靶向 CDC6。 细胞生理学杂志。 2019;234(6):9105–9117。 doi:10.1002/jcp.27587
10. 张明,王勇,蒋丽,等。 LncRNA CBR3-AS1 作为竞争性内源性 RNA 通过 JNK1/MEK4 介导的 MAPK 信号通路调节乳腺癌药物敏感性。 实验临床癌症研究杂志。 2021;40(1)。 号码:10.1186/s13046-021-01844-7
12. 姚芳,黄X,谢Z,等。 LINC02418 通过竞争性海绵 miR-372-3p 上调 EPHA2,从而促进结直肠癌中的 5-Fu/DDP 化疗耐药。 致癌作用。 2022;43(9):895–907。 doi:10.1093/carcin/bgac065
13. Ye Y,Li H,Bian J,Wang L,Wang Y,Huang H。通过生物信息学分析探索雌激素非依赖性子宫体子宫内膜癌的预后相关生物标志物。 国际现代医学杂志。 2021;14:9067–9081。 doi:10.2147/IJGM.S341345
14. 孙Z,麦华,薛C,等。 受 METTL3 调节的 Hsa-LINC02418/mmu-4930573I07Rik 通过增强 Trim21 介导的 PD-L1 泛素化来决定抗 PD-L1 免疫治疗功效。 癌症免疫治疗杂志。 2023;11(12):e007415。 doi:10.1136/jitc-2023-007415
15. Wang T, Zhai R, Lv X, Wang K, Xu J. LINC02418 通过海绵 miR-4677-3p 上调 KNL1 表达促进肺腺癌细胞的恶性行为。 BMC 脉医学。 2020;20(1):217。 号码:10.1186/s12890-020-01229-0
16. 田杰,崔鹏,李Y,等。 LINC02418 通过与 miR-34b-5p/BCL2 轴相互作用抑制细胞凋亡,从而促进结肠癌进展。 癌细胞国际。 2020;20:460。 号码:10.1186/s12935-020-01530-2
17. Rebello Alves LN、Dummer Meira D、Poppe Merigueti L 等。 乳腺癌的生物标志物:一个古老的故事,一个新的结局。 基因。 2023;14(7)。 doi:10.3390/genes14071364
18. Sung H、Ferlay J、Siegel RL 等。 2020 年全球癌症统计数据:GLOBOCAN 对全球 185 个国家 36 种癌症的发病率和死亡率的估计。 作为癌症杂志诊所。 2021;71(3):209–249。 doi:10.3322/caac.21660
19. Hosonaga M,Saya H,Arima Y。乳腺癌脑转移的分子和细胞机制。 癌症转移评论。 2020;39(3):711–720。 doi:10.1007/s10555-020-09881-y
20. Michaels E、Worthington RO、Rusiecki J。乳腺癌风险评估、筛查和一级预防。 北医学临床中心。 2023;107(2):271–284。 doi:10.1016/j.mcna.2022.10.007
21. 阿尔萨布·何. 长非编码 (lnc) RNA 在上皮间质转化 (EMT) 过程中 Wnt/β-catenin 信号通路调节中的病理作用。 病理学研究实践。 2023;248。
22. 于修国 JZ,亚光 H,何 Y。长非编码 RNA PVT1 通过 miR-30d-5p/RUNX2 轴在人结肠癌中发挥癌基因的作用。 J 好。 2018;23(1):48–54。
23. Li Y,Zeng Q,Qiu J,Pang T,Xian J,Zhang X.长非编码RNA UCA1通过抑制miR-206上调PTP1B表达促进乳腺癌。 癌细胞国际。 2019;19:275。 doi:10.1186/s12935-019-0958-z
24. 蒋X,周Y,孙AJ,薛JL。 NEAT1 通过调节 miR-448 和 ZEB1 促进乳腺癌进展。 细胞生理学杂志。 2018;233(11):8558–8566。 doi:10.1002/jcp.26470
25. 董浩,胡静,邹坤,等。 H3K27 乙酰化激活 LncRNA TINCR 通过靶向乳腺癌中的 MicroRNA-125b 促进曲妥珠单抗耐药和上皮间质转化。 摩尔癌。 2019;18(1):3。 号码:10.1186/s12943-018-0931-9
26. Lv M, Mao Q, Li J, Qiao J, Chen X, Luo S. LINC00665 的敲低通过 miR-3619-5p 的竞争性结合和连环蛋白 beta 1 的抑制抑制乳腺癌的增殖和侵袭。 细胞分子生物学快报。 2020; 25:43。 号码:10.1186/s11658-020-00235-8
27. Wu X, Kong C, Wu Y. 长基因间非蛋白编码 RNA 1094 (LINC01094) 通过调节 microRNA-340-5p (miR-340-5p)/E2F 转录因子 3 促进乳腺癌 (BC) 的进展 ( E2F3) 轴。 生物工程。 2021;12(1):9046–9057。 号码:10.1080/21655979.2021.1993715
28. HAN B. LncRNA LINC02418通过调节miR-4677-3p/SEC61G调节非小细胞肺癌细胞的增殖和凋亡。 欧洲医学评论药理学。 2019;23。
29. Qi BS L, Yang B, Yang B, Lu S. Su lu 线粒体 RNA 加工内切核糖核酸酶的长非编码 RNA 成分通过竞争性内源性 RNA 机制促进三阴性乳腺癌细胞的癌变。 乳腺癌杂志。 2021;24(5):428–442。 doi:10.4048/jbc.2021.24.e42
30. Bai Y,Zhang G,Cheng R,Yang R,Chu H。CASC15通过调节肺癌中的miR-766-5p/KLK12轴促进增殖和侵袭。 细胞周期。 2019;18(18):2323–2331。 号码:10.1080/15384101.2019.1646562
31. 黄超,苏旭,周德林,等。 诊断性和预测性lncRNA lnc-MPEG1-1通过占据miR-766-5p促进乳头状甲状腺癌细胞的增殖和转移。 摩尔热核酸。 2022;28:408–422。 doi:10.1016/j.omtn.2022.03.023
2024-04-24 10:35:14
1713956501
#LINC02418在乳腺癌中的表达及意义










